Viral diseases represent one of the greatest challenges for global aquaculture, triggering severe sanitary crises and millions in economic losses. Among these threats, infectious hematopoietic necrosis (IHN) stands out as a systemic and highly contagious pathology that severely impacts salmonids; consequently, its causative agent (IHNV) poses a paramount concern for salmon farmers worldwide.
Recently, a comprehensive scientific review published in the Journal of the World Aquaculture Society—conducted by researchers from the University of Tehran and Shahid Chamran University of Ahvaz, among other academic institutions—has exhaustively synthesized current knowledge on this pathogen, spanning its molecular structure and geographical distribution to its transmission mechanisms, host immune response, and contemporary vaccination strategies.
- 1 Study Key Points
- 2 What is IHNV, and how does it replicate?
- 3 Global distribution: From the United States to the world
- 4 IHNV Genotypes and Phylogeny: Keys to Designing Effective Vaccines
- 5 Clinical signs and internal pathology of IHNV
- 6 Immune response and protection phases against IHNV
- 7 Prevention, biosecurity, and vaccination strategies against IHNV
- 8 IHNV Diagnosis and Control: The Shield Before the Vaccine
- 9 Conclusion: Understanding the Virus to Protect Aquaculture Production
- 10 Entradas relacionadas:
Study Key Points
- The IHNV is a rhabdovirus possessing a single-stranded RNA genome that encodes six proteins, including the G-glycoprotein, which serves as the primary target for vaccine development.
- Horizontal transmission through water constitutes the main route of infection, with the gills serving as the pathogen’s most critical entry portal.
- Five major genogroups exist worldwide (U, M, L, E, and JRt/J), which strictly correlate with specific geographic regions.
- Diagnosis via viral isolation in cell lines (such as EPC or FHM) remains the ‘gold standard’ compared to rapid molecular techniques.
- DNA vaccines, commercialized in countries like Canada since 2005, provide robust and long-lasting protection that can extend up to two years.
What is IHNV, and how does it replicate?
The infectious hematopoietic necrosis virus (IHNV) belongs to the Rhabdoviridae family and is classified within the Novirhabdovirus genus. Its genetic material consists of a single-stranded, negative-sense, non-segmented RNA molecule approximately 11 kilobases in length. Despite this relatively simple molecular architecture, the pathogen possesses a formidable capacity to replicate rapidly, evade the immune system, and induce severe systemic damage in the host. The IHNV genome encodes six proteins essential for its biological cycle:
- Structural: The nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G), and RNA polymerase (L).
- Non-structural: The NV protein, unique to the Novirhabdovirus genus. The latter plays a crucial pathogenic role by inhibiting the host’s type I interferon production, thereby weakening the early immune response and promoting viral replication during the initial phases of infection.
IHNV replication occurs primarily within the cytoplasm of infected cells. Although the process can peak around the third day post-infection, viral kinetics are highly dependent on environmental temperature; for instance, in juvenile rainbow trout exposed to 15 °C, maximum viral titers are typically recorded between days 5 and 7.
Transmission mechanisms and species affected by IHNV
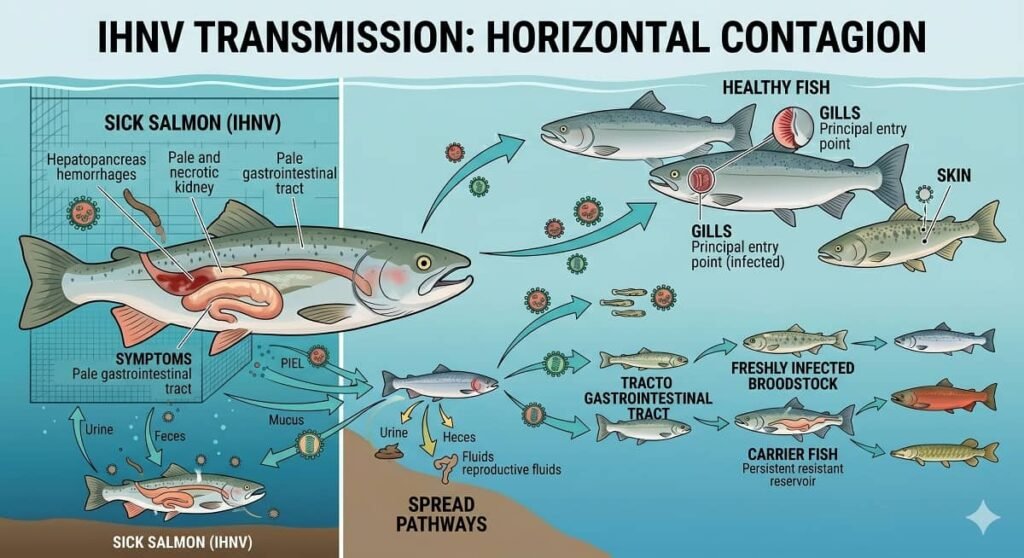
Horizontal transmission through water constitutes the primary pathway for IHNV dissemination, a phenomenon that intensifies within hatcheries and intensive aquaculture production systems. The pathogen sheds via urine, feces, mucus, and reproductive fluids from infected specimens, gaining entry into new hosts through the gills, skin, or gastrointestinal tract; consequently, under high-density rearing conditions, even minimal viral loads can trigger rapid and devastating epizootic outbreaks.
IHNV predominantly affects salmonids of the genera Oncorhynchus, Salmo, and Salvelinus, with the most susceptible species including:
- Rainbow trout (Oncorhynchus mykiss)
- Atlantic salmon (Salmo salar)
- Chinook, coho, and sockeye salmon
- Northern pike (Esox lucius), a non-salmonid species that has also demonstrated susceptibility to the virus.
Fry and juveniles represent the most vulnerable life stages, sustaining mortality rates that can reach up to 90%. Furthermore, specimens surviving these outbreaks frequently become asymptomatic reservoirs of the virus, perpetuating infection risks and compromising the biosecurity of subsequent production cycles.
Global distribution: From the United States to the world
The IHNV was first identified during the 1950s in sockeye salmon (Oncorhynchus nerka) hatcheries across the western United States; since then, its geographical expansion has closely mirrored international aquaculture trade routes. The transport of infected eggs and specimens propagated the pathogen first to other North American regions, subsequently to Asia (Japan, China, South Korea, and Iran), and ultimately to Europe, where it is now established in nations such as France, Germany, Italy, Spain, Denmark, and the Netherlands.
Due to its high impact, the World Organisation for Animal Health (WOAH) lists it officially as one of the three most critical rhabdoviruses affecting commercially important fish, while no cases of the disease have been reported in Africa or the Southern Hemisphere to date.
Stay Always Informed
Join our communities to instantly receive the most important news, reports, and analysis from the aquaculture industry.
IHNV Genotypes and Phylogeny: Keys to Designing Effective Vaccines
One of the most significant contributions of this scientific review is the phylogenetic analysis of diverse IHNV strains, evaluating the G, N, M, and NV genes through 206 global sequences retrieved from GenBank. The findings confirm that the virus organizes into distinct genotypes with a pronounced geographic distribution. Three groups predominate in North America: U (upper, spanning from Alaska to Oregon), M (middle, located in Idaho and the Columbia River basin, featuring the highly pathogenic ‘220-90’ strain), and L (lower, present in California and southern Oregon); meanwhile, genotype E encompasses European isolates, and Japan and South Korea harbor genotype J.
The most critical discovery for sanitary management in non-native production zones occurs in Iran, where local strains conclusively cluster within the European genotype E—specifically alongside Italian isolates—backed by an 89% bootstrap value. This finding demonstrates a historical introduction of the pathogen from regions prevalent in genotype E, likely linked to the importation of infected eggs or live specimens. The practical implication is immediate and vital for the industry: vaccines designed based on genotype E will yield superior immunological efficacy for Iranian salmon farming.
Clinical signs and internal pathology of IHNV
The manifestation of infectious hematopoietic necrosis varies according to the species, size, and life stage of the fish. In the most vulnerable phases (fry and juveniles weighing 0.2 to 8 grams), the acute presentation of the pathology triggers critical mortality rates of up to 90%.
External and Internal Symptomatology
Specimens affected by the acute form of IHN exhibit skin darkening, lethargy, pale gills due to severe anemia, and exophthalmia (popeye). A characteristic clinical sign is the presence of an opaque, mucous fecal pseudocast trailing from the rectum, accompanied by chevron-shaped hemorrhages along the lateral musculature, as well as petechiae in the oral cavity and on the body surface. Internally, liver and kidney pallor, ascites (fluid accumulation in the abdominal cavity), and reddish-yellow mucoid fluids within the stomach are observed. Petechial hemorrhages spread through the adipose tissue, swim bladder, peritoneum, and meninges.
Disease Variants and Histopathological Diagnosis
- Neurotropic IHN: Affects larger juveniles (over 8 grams), causing tetany, erratic swimming behavior, and spinal deformities such as scoliosis and lordosis.
- Epitheliotropic (Branchial) IHN: Identified in larger fish (50 to 100 grams). It induces no systemic symptoms but causes severe respiratory distress that compromises feed conversion ratios and long-term growth.
At the histopathological level, the virus induces extensive necrosis in the hematopoietic tissues of the posterior kidney and spleen, as well as in the pancreas and digestive system. Necrosis of the granular cells in the digestive tract establishes itself as a classic pathognomonic finding, visible exclusively in fish older than 3 to 4 months.
Immune response and protection phases against IHNV
Against IHNV infection, salmonids deploy an immune response structured into three consecutive temporal phases:
- Early Antiviral Response (EAVR): Activated within hours or days of pathogen exposure, this mechanism stimulates antiviral proteins—such as interferon-induced Mx—conferring a non-specific protection that extends for 3 to 4 weeks.
- Specific Antiviral Response (SAVR): Consolidating between the third and fourth weeks post-infection, it involves the production of neutralizing antibodies primarily directed against the G-glycoprotein, sustaining host protection for several months.
- Long-Term Antiviral Response (LAVR): This immunological memory phase can persist for over 25 months in rainbow trout, although a gradual decline in circulating antibody levels is observed.
This immune progression underscores why the G-glycoprotein remains the cornerstone for biotechnological vaccine development, representing the critical antigen capable of triggering the most robust and long-lasting neutralizing response.
Prevention, biosecurity, and vaccination strategies against IHNV
Effective management of the infectious hematopoietic necrosis virus demands a strict preventive approach, given that no curative treatments exist once systemic infection is declared within fish populations.
Disinfection and Biosecurity Measures
Safe production initiates with sourcing eggs and fry from certified virus-free broodstock reared in specific pathogen-free (SPF) water facilities; when the sanitary status of the eggs is uncertain, mandatory surface disinfection using iodophor-based solutions is prescribed.
Experimental trials demonstrate that iodophor doses of 100 mg/L achieve a 99.98% viral eradication rate on the egg surface. Additionally, for feed management, dehydration or thermal sterilization at 60 °C for 30 minutes is recommended to ensure the complete inactivation of any circulating viral particles.
Furthermore, the administration of bacterial probiotics (such as Pseudomonas sp., Vibrio sp., Lactobacillus, and Pediococcus acidilactici) has emerged as a sustainable alternative in biosecurity strategies, as certain bacterial strains isolated from fish farms release extracellular metabolites with proven antiviral properties capable of reducing IHNV plaque formation by over 50% in laboratory assays.
Advances in IHNV Vaccines
The scientific review analyzes the development and efficacy of four vaccine categories:
- Inactivated Vaccines: Based on formalin-inactivated viruses and administered via intraperitoneal injection, they demonstrate efficacy in larger fish before their transfer to marine cages; however, immersion immunization with non-replicating antigens remains insufficient for large-scale disease containment.
- Subunit Vaccines: Formulated from recombinant G-proteins expressed in various heterologous systems (bacteria, yeast, or insect cells), these formulations have failed to confer significant protection to date, and their mass production for fry presents a severe economic hurdle.
- DNA Vaccines: Consolidating as the most effective option available, the first of these—named APEX-IHN (Novartis Animal Health Canada Inc.)—was licensed in Canada in 2005 for Atlantic salmon. Administered intramuscularly at doses of 0.1 to 1 µg, they induce robust immunity in juvenile rainbow trout and protect specimens for up to two years; nonetheless, no commercial vaccine against IHNV is authorized for use in Europe yet.
- Recombinant Live Vaccines: Genetic engineering of the IHNV genome has enabled the development of attenuated viral vectors capable of reducing pathogenicity while expressing antigens from other viruses (such as VHSV, IPNV, or ISAV), achieving simultaneous protection against multiple pathologies and recording post-vaccination mortality rates of a mere 10%.
IHNV Diagnosis and Control: The Shield Before the Vaccine
Viral isolation in cell cultures remains the gold standard for IHNV diagnosis, although molecular techniques such as RT-qPCR offer greater speed and analytical sensitivity, requiring EPC or FHM cell lines incubated at 15 °C for 7 to 10 days.
Within the scope of preventive management, key guidelines to mitigate the pathogen’s impact include utilizing certified virus-free eggs, applying iodophor solutions at 100 mg/L to achieve a 99.98% surface eradication rate, disinfecting water inflows via UV radiation or ozone, and implementing strict biosecurity protocols alongside probiotics like Lactobacillus, Pseudomonas, and Pediococcus acidilactici to help reduce viral loads during active outbreaks.
Conclusion: Understanding the Virus to Protect Aquaculture Production
This comprehensive review of IHNV conclusively demonstrates that understanding the pathogen’s biology, epidemiology, and phylogeny transcends purely academic interest; instead, it establishes itself as an indispensable strategic tool for optimizing decision-making within the global aquaculture industry.
On one hand, precisely determining the circulating genotype in a specific region represents the critical factor for selecting the most appropriate vaccine variant; on the other hand, rigorously identifying transmission pathways enables the design of significantly more effective biosecurity protocols in farming facilities. Ultimately, accelerating the development and regulation of accessible DNA vaccines could mark a historic milestone for salmon farming across strategic regions such as Europe, Asia, and Latin America.
Contact
Hooman Rahmati-Holasoo
Department of Aquatic Animal Health, Faculty of Veterinary Medicine, University of Tehran
Tehran, Iran.
Email: rahmatih@ut.ac.ir
Reference (open access)
Dezfuly, Z. T., Marandi, A., Abbasi, M., Bashiri, M., Ghalyanchilangeroudi, A., Kafi, Z. Z., Zorriehzahra, M. J., Aghaiyan, L., Jørgensen, G., & Rahmati-Holasoo, H. (2026). Infectious hematopoietic necrosis virus (IHNV) in fish and its consequences for fish farming: A comprehensive review. Journal of the World Aquaculture Society, 57(3), e70114. https://doi.org/10.1111/jwas.70114
Editor at the digital magazine AquaHoy. He holds a degree in Aquaculture Biology from the National University of Santa (UNS) and a Master’s degree in Science and Innovation Management from the Polytechnic University of Valencia, with postgraduate diplomas in Business Innovation and Innovation Management. He possesses extensive experience in the aquaculture and fisheries sector, having led the Fisheries Innovation Unit of the National Program for Innovation in Fisheries and Aquaculture (PNIPA). He has served as a senior consultant in technology watch, an innovation project formulator and advisor, and a lecturer at UNS. He is a member of the Peruvian College of Biologists and was recognized by the World Aquaculture Society (WAS) in 2016 for his contribution to aquaculture.
: Technology, Design, and Profitability")
 2026: A Comprehensive Guide to Aquaculture 4.0 via AI and IoT Integration")




